
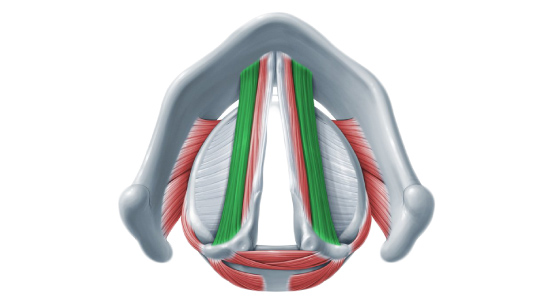
La especialización del músculo tiroaritenoideo
El tiroaritenoideo (TA) se describe como el músculo más importante para la fonación (Han, Wang, Fischman, Biller, & Sanders, 1999; Périé, Agbulut, Lacau St Guily, & Butler-Browne, 2000; Hoh, 2005). Se extiende desde la superficie interna del cartílago tiroides hasta la superficie anterior de la apófisis vocal. Se trata de un músculo que presenta dos grandes porciones, una denominada muscularis (porción lateral) y otra, vocalis (porción medial). Se ha estimado que la primera se relaciona con los movimientos de aducción en actividades como la tos, mientras que la segunda, con la fonación (Han et al., 1999). Figura 1. En izquierda se observa la rama vocalis, mientras que a la derecha, la muscularis
Figura 1. En izquierda se observa la rama vocalis, mientras que a la derecha, la muscularis
En los seres humanos, las distintas necesidades comunicativas que su laringe debe satisfacer han llevado a que este músculo se adapte y adquiera diferenciaciones histoquímicas importantes con respecto a otros mamíferos (Wu, Crumley, Armstrong, & Caiozzo, 2000).
Dentro de estas adaptaciones, la porción muscularis es la de mayor similitud con respecto a otros mamíferos. En su interior, presenta fibras veloces y fuertes (Wu et al., 2000), tales como las IIX (López, Zárate, Cantín, Zavando, & Suazo Galdames, 2011) e isoformas de cadena pesada de miosina (MyHC) del tipo L y 2B. Este tipo de configuración de fibras y de MyHC existe cuando se necesitan contracciones ultra rápidas y de corta duración, tal como las que se generan cuando se activa el reflejo protector de la vía aérea (Tellis, Rosen, Thekdi, & Sciote, 2004).
En la porción vocalis, donde las adaptaciones son altamente notorias en comparación al TA de otros mamíferos, se ha observado una importante cantidad de fibras tónicas lentas (Han et al., 1999) y tónicas (López et al., 2011). A su vez, también se ha descrito la presencia de fibras tipo IIA o MyHC IIA (Shiotani, Westra, & Flint, 1999; Wu et al., 2000; Li, Lehar, Nakagawa, Hoh, & Flint, 2004).
Se ha estimado que la resistencia superior que entregan las fibras tónicas lentas, tónicas y tipo IIA permite que la rama vocalis participe activamente en el control de la duración de la duración, dándole así, un grado de fatigabilidad superior a otros músculos del cuerpo humano (Han et al., 1999).
Cuando la fonación se realiza con intensidades y tonalidades consideradas como conversacionales, serán principalmente las fibras tónicas y tónicas lentas las que participan activamente (Han et al., 1999). Si los requerimientos aumentan y se solicita mayor intensidad vocal, serían las fibras glucolíticas/mixtas las que se recluten y complementen el trabajo de las fibras oxidativas (Sandage & Smith, 2017). Con respecto al control de los tonos que se ejecutan con preponderancia del mecanismo tiroaritenoideo, se ha teorizado que el músculo vocalis complementaría sus funciones con las fibras tipo I/IIA del cricotiroideo (Shiotani et al., 1999) y con las fibras oxidativas e híbridas del interaritenoideo (Tellis et al., 2004).
De modo general, la porción vocalis presenta una mezcla de fibras que permiten generar la fuerza y la resistencia vocal que se requiere en la vida diaria (Talbot & Maves, 2016). No obstante, gracias a su entrenamiento, se pueden alcanzar adaptaciones que favorezcan el desarrollo particular de alguna de estas propiedades, tal como sucede con los deportistas (Serrano et al., 2019). Estas serán estudiadas en artículos posteriores.
El músculo TA no solo entrega particularidades en su distribución de fibras y de cadenas pesadas de miosina, sino que aspectos como su propiocepción también son específicos en su morfología. Los receptores sensoriales de los músculos cumplen un importante papel durante la contracción, gracias a ellos, el músculo produce la fuerza y velocidad de contracción precisa para la función solicitada (Thomas, Harrison, & Stemple, 2008).
El huso muscular es un receptor sensorial que se ubica en el interior del músculo. Se caracteriza porque detecta sus cambios de longitud (mide la longitud o cambio de estiramiento del músculo). Debido a su función, este receptor le proporciona la información necesaria al sistema nervioso para proteger al músculo ante un cambio de longitud brusco o excesivo (López Chicharro & Fernández Vaquero, 2006). En el músculo tiroaritenoideo se han observado una densa población de husos neuromusculares en la porción vocalis. Se ha indicado que su presencia permite que el sistema nervioso sea capaz se detectar y monitorizar los cambios de elongación que tiene el músculo a medida se asciende en la escala musical (Sanders et al., 1998).
Adicionalmente, se ha observado la presencia de propioceptores en la mucosa de los pliegues vocales. Estos tendrían la función de enviar la información necesaria para la activación muscular durante la aducción (Andreatta, Mann, Poletto, & Ludlow, 2002).
Los puntos analizados en este artículo son solo algunas de las particularidades que tiene el músculo tiroaritenoideo (tabla 1). A pesar de no ser desarrolladas, es también necesario considerar la importante densidad mitocrondrial, los pequeños depósitos de grasa y la existencia de mioglobinas que hay en su interior.
 Tabla 1. Características del músculo tiroaritenoideo (Thomas et al., 2008)
Tabla 1. Características del músculo tiroaritenoideo (Thomas et al., 2008)
Conclusión
El tiroaritenoideo es el principal músculo de la fonación, en su interior presenta dos porciones, una denominada muscularis y otra, vocalis. En su porción muscularis se encuentran, primordialmente, fibras ultra rápidas, mientras que en su porción vocalis se localizan fibras tónicas, tónicas lentas y de tipo IIA. Estas últimas son parte de las adaptaciones evolutivas que ha sufrido el músculo para cumplir con las demandas fonatorias propias del ser humano.
Bibliografía
Andreatta, R. D., Mann, E. A., Poletto, C. J., & Ludlow, C. L. (2002). Mucosal afferents mediate laryngeal adductor responses in the cat. Journal of Applied Physiology, 93(5), 1622–1629. https://doi.org/10.1152/japplphysiol.00417.2002
Asanau, A., Timoshenko, A. P., Prades, J. M., Galusca, B., Martin, C., & Féasson, L. (2011). Posterior cricoarytenoid bellies: Relationship between their function and histology. Journal of Voice, 25(2). https://doi.org/10.1016/j.jvoice.2010.11.004
Han, Y., Wang, J., Fischman, D. A., Biller, H. F., & Sanders, I. (1999). Slow tonic muscle fibers in the thyroarytenoid muscles of human vocal folds; a possible specialization for speech. Anatomical Record, 256(2), 146–157. https://doi.org/10.1002/(SICI)1097-0185(19991001)256:2<146::AID-AR5>3.0.CO;2-8
Hoh, J. F. Y. (2005, February). Laryngeal muscle fibre types. Acta Physiologica Scandinavica, Vol. 183, pp. 133–149. https://doi.org/10.1111/j.1365-201X.2004.01402.x
Li, Z., Lehar, H., Nakagawa, H., Hoh, J., & Flint, P. (2004). Differential expression of myosin heavy chain isoforms between abductor and adductor muscles in the human larynx. Otolaryngol Head Neck Surg, 130(2), 217–222.
López Chicharro, J., & Fernández Vaquero, A. (2006). Fisiología del ejercicio. Médica Panamericana.
López, M. J., Zárate, M., Cantín, M., Zavando, D., & Suazo Galdames, I. (2011). Immunohistochemical Fiber Type Profile in the Human Vocal Muscle. International Journal of Morphology, 29(4), 1158–1161. https://doi.org/10.4067/s0717-95022011000400014
Périé, S., Agbulut, O., Lacau St Guily, J., & Butler-Browne, G. S. (2000). Myosin heavy chain expression in human laryngeal muscle fibers: A biochemical study. Annals of Otology, Rhinology and Laryngology, 109(2), 216–220. https://doi.org/10.1177/000348940010900218
Sandage, M. J., & Smith, A. G. (2017). Muscle Bioenergetic Considerations for Intrinsic Laryngeal Skeletal Muscle Physiology. Journal of Speech, Language, and Hearing Research, 60(5), 1254–1263. https://doi.org/10.1044/2016_JSLHR-S-16-0192
Sanders, I., Han, Y., Wang, J., & Biller, H. (1998). Muscle spindles are concentrated in the superior vocalis subcompartment of the human thyroarytenoid muscle. Journal of Voice, 12(1), 7–16. https://doi.org/10.1016/S0892-1997(98)80070-2
Sciote, J. J., Morris, T. J., Horton, M. J., Brandon, C. A., & Rosen, C. (2002). Unloaded shortening velocity and myosin heavy chain variations in human laryngeal muscle fibers. Annals of Otology, Rhinology and Laryngology, 111(2), 120–127. https://doi.org/10.1177/000348940211100203
Serrano, N., Colenso-Semple, L. M., Lazauskus, K. K., Siu, J. W., Bagley, J. R., Lockie, R. G., … Galpin, A. J. (2019). Extraordinary fast-twitch fiber abundance in elite weightlifters. PloS One, 14(3), e0207975. https://doi.org/10.1371/journal.pone.0207975
Shiotani, A., Westra, W. H., & Flint, P. W. (1999). Myosin heavy chain composition in human laryngeal muscles. Laryngoscope, 109(9), 1521–1524. https://doi.org/10.1097/00005537-199909000-00030
Smerdu, V., & Cvetko, E. (2013). Myosin heavy chain-2b transcripts and isoform are expressed in human laryngeal muscles. Cells Tissues Organs, 198(1), 75–86. https://doi.org/10.1159/000351293
Talbot, J., & Maves, L. (2016). Skeletal muscle fiber type: using insights from muscle developmental biology to dissect targets for susceptibility and resistance to muscle disease. Wiley Interdisciplinary Reviews. Developmental Biology, 5(4), 518–534. https://doi.org/10.1002/wdev.230
Tellis, C. M., Rosen, C., Thekdi, A., & Sciote, J. J. (2004). Anatomy and fiber type composition of human interarytenoid muscle. Annals of Otology, Rhinology and Laryngology, 113(2), 97–107. https://doi.org/10.1177/000348940411300203
Thomas, L. B., Harrison, A. L., & Stemple, J. C. (2008, July). Aging Thyroarytenoid and Limb Skeletal Muscle: Lessons in Contrast. Journal of Voice, Vol. 22, pp. 430–450. https://doi.org/10.1016/j.jvoice.2006.11.006
Wu, Y. Z., Crumley, R. L., Armstrong, W. B., & Caiozzo, V. J. (2000). New perspectives about human laryngeal muscle: Single-fiber analyses and interspecies comparisons. Archives of Otolaryngology – Head and Neck Surgery, 126(7), 857–864. https://doi.org/10.1001/archotol.126.7.857